





One of Darwin’s most important contributions made to the science of evolution is the concept of natural selection. Despite its widespread unacceptance for several decades, the idea that members of a species compete with each other for resources and that individuals who are better adapted to their lifestyle have a better chance of surviving to reproduce was revolutionary to the field of evolution. Today, natural selection forms the basis for our understanding of how species change over time.
In our discussions of Darwin’s theories, we looked at Darwin's proposed mechanism for evolution, natural selection. It had five postulates:
-
Individuals are variable.
-
Some variations are passed down.
-
More offspring are produced than can survive.
-
Survival and reproduction are not random.
-
The history of Earth is long.
This last point was proven by geologists and astronomers.
Individuals Are Variable
Darwin knew that individuals were variable, that is, each individual in a population carried a unique set of traits. What he did not know is what produced this variability, namely genetic differences. Variation in the genes of individuals arises from several sources. Mutation, the alteration of existing genes to form new alleles, can arise from copying errors during DNA replication, DNA damage, and repair or recombination during cell division. Variation also arises from sexual reproduction; wherein new combinations of DNA are created through the independent assortment of genes.
Some Variations Are Passed Down
This statement was a truly unique portion of Darwin's theory. In 1856, he did not know about DNA. He did not know about recombination events. He did not even know about genes. He merely understood that for selection to occur, variations must be transmittable from parent to offspring. We now know that variation is caused by differences in genes, and that genes are passed on to offspring. More importantly, different genes are passed on to offspring independently of each other (independent assortment).
More Offspring Are Produced than Can Survive
In most generations, more offspring are born than can survive to reproductive age given selection pressures such as predation and limited food supply. For example, many fish lay hundreds or even thousands of eggs at once, yet most of the young will be eaten or will starve before they can produce young of their own and pass down their genes (and the genes of their parents).
Reproduction and Survival Are Not Random
This portion of Darwin's theory is what we know as "survival of the fittest." Since more offspring are produced than can survive, some must die. Those that survive are those that have the greatest fitness. A trait that increases an individual's fitness is called an adaptation. Those individuals with the genes that convey traits that are best adapted to the environment in which the organism lives (those with high fitness) are more likely to survive and reproduce than those that are less adapted to their environment (those with low fitness).
On the genetic level, a specific allele for a trait can produce an adaptation and convey greater fitness. An individual with greater fitness is more likely to reproduce and pass this allele on to the next generation. Since more fit individuals produce more offspring, the percentage of individuals in the next generation with the fit allele—the allelic frequency of that allele—will increase. As this process repeats itself over many generations, evolution occurs, since the beneficial allele comes to exist within most of the population.
Selective Pressures
Natural selection may act to change a trait in many different ways. When selection pressures favor the average form of the trait, selection is said to be stabilizing. Directional selection occurs when selection pressures favor one extreme of the trait distribution. Selection is disruptive when the average form of the trait is selected against while either extreme is favored.
In addition to natural selection, there are two other types of selection. Sexual selection, which Darwin believed was distinct from natural selection, involves the selection of traits based on their role in courtship and mating. Artificial selection is the selective breeding of species by humans to increase desirable traits, though the traits do not necessarily have to confer greater fitness.
Natural selection can take many forms. To make talking about this easier, we will consider the distribution of traits across a population in graphical form. In Figure 7.01 we see the normal bell curve of trait distribution. For example, if we were talking about height as a trait, we would see that without any selection pressure on this trait, the heights of individuals in a population would vary, with most individuals being of an average height and fewer being extremely short or extremely tall. However, when selection pressures act on a trait, this distribution can be altered.
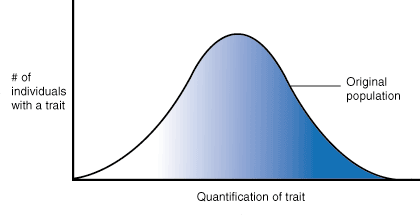
Figure 7.01: The standard distribution of a trait across a population
Stabilizing Selection
When selective pressures select against the two extremes of a trait, the population experiences stabilizing selection. For example, plant height might be acted on by stabilizing selection. A plant that is too short may not be able to compete with other plants for sunlight. However, extremely tall plants may be more susceptible to wind damage. Combined, these two selection pressures select to maintain plants of medium height. The number of plants of medium height will increase while the numbers of short and tall plants will decrease.
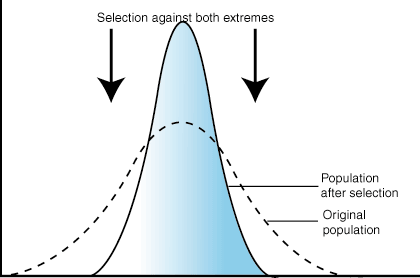
Figure 7.02: The effect of stabilizing selection on trait distribution
Directional Selection
In directional selection, one extreme of the trait distribution experiences selection against it. The result is that the population's trait distribution shifts toward the other extreme. In the case of such selection, the mean of the population graph shifts. Using the familiar example of giraffe necks, there was a selection pressure against short necks, since individuals with short necks could not reach as many leaves on which to feed. As a result, the distribution of neck length shifted to favor individuals with long necks.
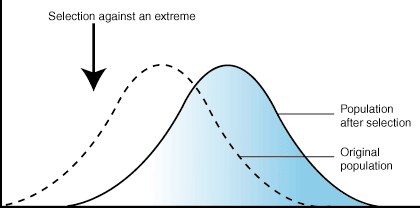
Figure 7.03: The effect of directional selection on trait distribution
Disruptive Selection
In disruptive selection, selection pressures act against individuals in the middle of the trait distribution. The result is a bimodal, or two-peaked, curve in which the two extremes of the curve create their own smaller curves. For example, imagine a plant of extremely variable height that is pollinated by three different pollinators: one that is attracted to short plants, another prefers plants of medium height, and a third that visits only the tallest plants. If the pollinator that prefers plants of medium height disappeared from an area, medium-height plants would be selected against and the population would tend toward both short and tall, but not medium-height plants. Such a population, in which multiple distinct forms or morphs exist is said to be polymorphic.
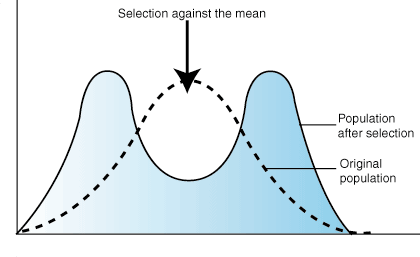
Figure 7.04: The effect of disruptive selection on trait distribution



