




Secondary growth is a growth of thickness. Secondary growth is a product of two different, though related, tissues, which both fall under the umbrella term lateral meristems. Vascular cambium exists between xylem and phloem; on its inside the cambium produces what is known as secondary xylem, on its outside it forms secondary phloem. The primary xylem and phloem are pushed further inward and outward, respectively. As xylem gets older it often becomes clogged and ceases to function; this tissue is called heartwood, in direct contrast to sapwood, which comprises the functioning xylem cells. The vascular cambium is more productive during the growing seasons. During the rest of the year it creates little growth. This phenomenon creates distinct rings of growth, each ring representing a single growing season. By studying these rings, it is possible to calculate the age of a plant, and even possible to determine the specific conditions of a particular growing season. The second lateral meristem is called cork cambium, and is responsible for the formation of cork (bark), which replaces the epidermis is the protective covering of shoot and root.
Gymnosperms
Gymnosperms are commonly known as conifers, and includes spruces, firs, hemlocks, and othercommon evergreens. Like all tracheophytes, gymnosperms contain vascular tissues. They have developed seeds and nonflagellated sperm; male gametes (carried inside pollen grains) are moved by the wind instead of through water.
The dominant phase in the gymnosperm life cycle is the
diploid (sporophyte) stage; the gametophytes are very small and
cannot exist independent of the parent plant. Male and female cones, the
reproductive structures of the sporophyte, produce two different kinds of
haploid spores: microspores (male) and megaspores (female).
This phenomenon is called heterospory. These spores give rise to
gametophytes of the same sex, which in turn produce the gametes.
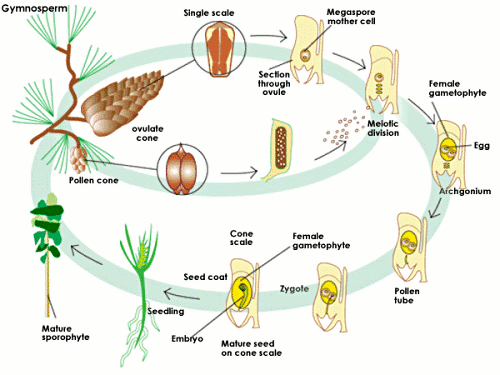
Gymnosperm are also characterized by a specialized fertilization process, involving differentiated male and female gametophytes. Fertilization occurs when pollen grains (male gametophytes) are carried by the wind to the open end of an ovule, which contains the eggs, or female gametophyte. For a (much) more detailed exploration of this process, see gymnosperm fertilization
Angiosperms
Angiosperms are typically divided into two classes: monocots (including grasses, grains, and spring-flowering bulbs such as daffodils) and dicots (including oaks, elms, sunflowers, and roses). As discussed in Xylem and Phloem , angiosperms have a vascular advantage over gymnosperms. The vessel elements in their vascular tissue, which evolved from the tracheids found in conifers, are more specialized for conducting fluids. In addition, fibers within angiosperm xylem give added support to the plant structure. Another positive adaptation that is exclusive to angiosperms is the flower, which attracts insects and thus facilitates the transfer of pollen. Flowers, the reproductive structures of angiosperms, take the place of gymnosperm cones. Furthermore, while the ovules of gymnosperms are exposed on the surface of the cone), angiosperm seeds (which develop from ovules) are enclosed within an ovary. This ovary later matures into a fruit, which aids in the dispersal of the seeds (through animals or wind) and protects the seeds from drying out.
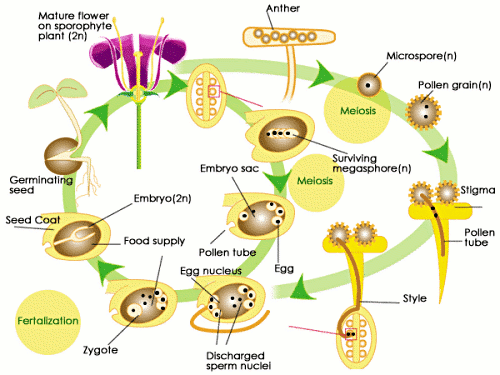
Much of the angiosperm life cycle resembles that of gymnosperms. The sporophyte stage dominates, and the gametophytes are even smaller than those of gymnosperms. The mature diploid plant produces male and female haploid spores through heterospory, which gives rise to single-sex gametophytes, which in term produce gametes. These gametes, through either self-pollination or cross-pollination, join to form a diploid zygote that eventually becomes a seed for a new angiosperm. For more information on the life cycles of angiosperms see Alternation of Generations.



